起源
拉马克的用进废退理论是研究进化的开端。
之后便是达尔文的自然选择。
两位伟人都告诉我们:进化很重要。那么如何研究进化呢?一种是看死的(化石),一种是看活的(比较形态学、比较解剖学)。
两种传统方法都有一定的局限性,而今天我们研究的是分子进化,即DNA、RNA以及蛋白质序列上的进化关系,利用软件从分子水平上构建物种的进化树。
当然,分子进化有两个基本的假设条件,只有接受这两个假设,分子进化理论才得以实施。
- DNA、RNA或蛋白质序列包含了物种的所有进化史信息。
- 分子钟理论。即一个特定基因或蛋白质的进化变异速度在不同物种中是基本恒定的。
同源
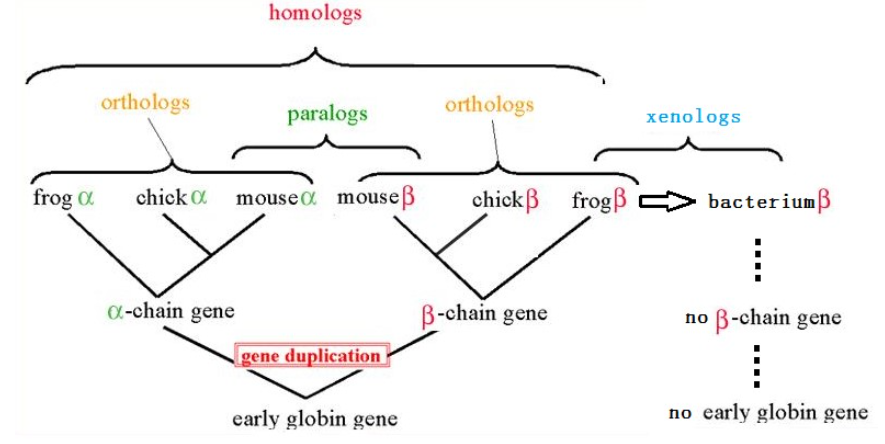
同源,即来源相同。这是一个论断,即来源于共同祖先的相似序列为同源序列。否则是不同源的。
同源又分为三种:直系同源、旁系同源和异同源。
直系同源(Orthologs):来自于不同物种的由垂直家系,也就是物种形成,进化而来的基因,并且典型的保留与原始基因相同的功能。也就是说,随着进化分支,一个基因进入了不同的物种,并保留了原有功能。这时,不同物种中的这个基因就属于直系同源。
旁系同源(Paralogs):在同一物种中的来源于基因复制的基因,可能会进化出新的但与原功能相关的功能来。基因复制产生了两个重复的基因,多出来的这个有几种命运,一个是又丢了。复制出来发现没有用,又删了。另一种命运是演化出了新的功能。如果这个新功能是往好的方向发展,就会被保留下了,如果是往不好的方面发展,就会被自然选择淘汰。还有一种命运,就是被放置不用。复制出来以后,又加了个终止子,既不表达,也不删除,搁那里搁着不管,成了伪基因。被保留下来的具有新功能的基因与另一个复制出来的基因之间就是旁系同源。
异同源(Xenologs):通过水平基因转移,来源于共生或病毒侵染所产生的相似基因。异同源的产生不是垂直进化而来的,也不是平行复制产生的,而是由于原核生物与真核生物的接触,比如病毒感染,在跨度巨大的物种间跳跃转移产生的。
系统发生树
研究分子进化所要构建的系统发生树,也叫分子树。
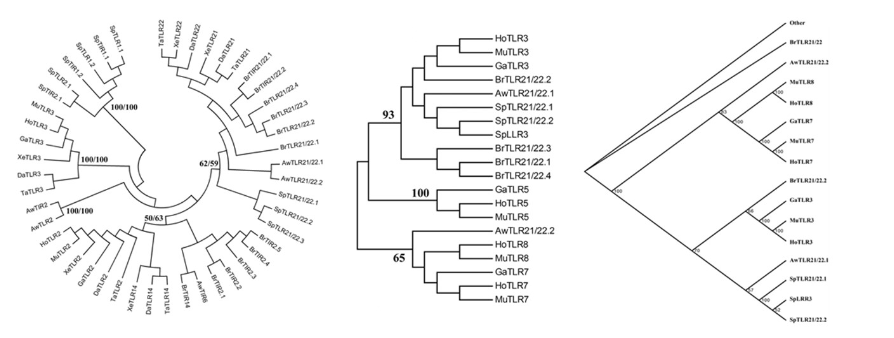
种类:有根树、无根树。
在线软件:
 http://evolution.genetics.washington.edu/phylip.html
http://evolution.genetics.washington.edu/phylip.html http://www.megasoftware.net
http://www.megasoftware.net http://paup.csit.fsu.edu/
http://paup.csit.fsu.edu/ http://www.atgc-montpellier.fr/phyml/
http://www.atgc-montpellier.fr/phyml/ http://mrbayes.csit.fsu.edu/
http://mrbayes.csit.fsu.edu/